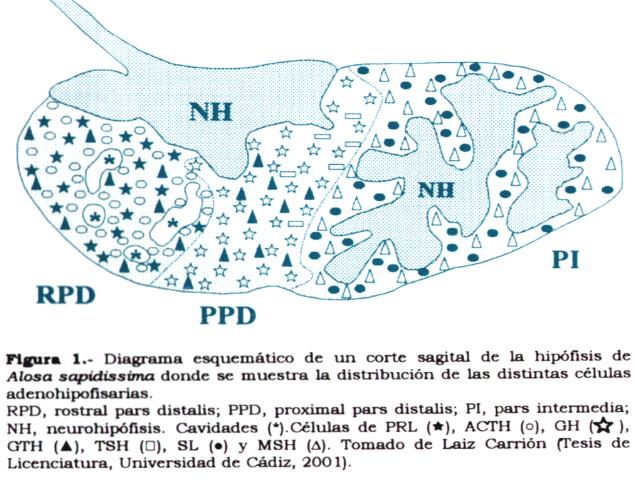
La adenohipófisis de los teleósteos
La hipófisis es una glándula clave para el control hormonal de diferentes funciones, tales como reproducción, desarrollo, crecimiento, osmorre-gulación, metabolismo, etc. Esta glándula es un lugar de síntesis, almacenaje y liberación de varias hormonas peptídicas que actúan sobre estos procesos. De este modo, la hipófisis puede ser considerada como un transductor que gracias a sus secreciones permite que el sistema nervioso central controle un rango muy amplio de funciones endocrinas.
Esta glándula se compone de dos tipos de tejidos con orígenes distintos: i) la adenohipófisis, originada por una evaginación ectodérmica del techo de la cavidad bucal embrionaria, y ii) la neurohipófisis de origen nervioso, originada de una invaginación del proceso ventral del diencéfalo. Los tipos celulares de la adenohipófisis se encuentran localizados en determinadas partes de la glándula, de manera que dos o más tipos celulares constituyen un lóbulo hipofisario. Las células que integran los diferentes lóbulos hipofisarios se distinguen por su morfología, afinidades tintoriales, reacciones inmunocitoquímicas y por su función fisiológica. Desde el punto de vista anatómico, la adenohipófisis de peces teleósteos presenta tres partes diferenciadas:
i) Pars distalis rostralis (PDR), en la posición más rostral de la glándula. En ella se localizan las células lactotropas o productoras de la hormona lactotropa (prolactina, PRL), y las células corticotropas o productoras de la hormona estimulante de las cápsulas adrenales (adrenocorticotropina, ACTH).
ii) Pars distalis proximalis (PDP), en posición caudal a la PDR. Contiene las células somatotropas o sintetizadoras de la hormona del crecimiento (somatotropina, GH), células tirotropas o productoras de la hormona estimulante del tiroides (tirotropina, TSH), y células gonadotropas o sintetizadoras de las hormonas estimulantes de las gónadas (gonadotropinas, GTH I y GTH II).
iii) Pars intermedia (PI), localizada en la parte más caudal de la glándula. En ella se hallan las células melanotropas o productoras de la hormona estimulante de los melanocitos (melanotropina, MSH), y las células somatolactotropas o sintetizadoras de la hormona somatolactina (SL).
La distribución de hormonas adenohipofisarias puede variar ligeramente entre especies, aunque la distribución más común es la explicada anteriormente. En algunos teleósteos es posible observar células GTH en la PDR y PI (ver figura 1).
Desarrollo de la adenohipófisis
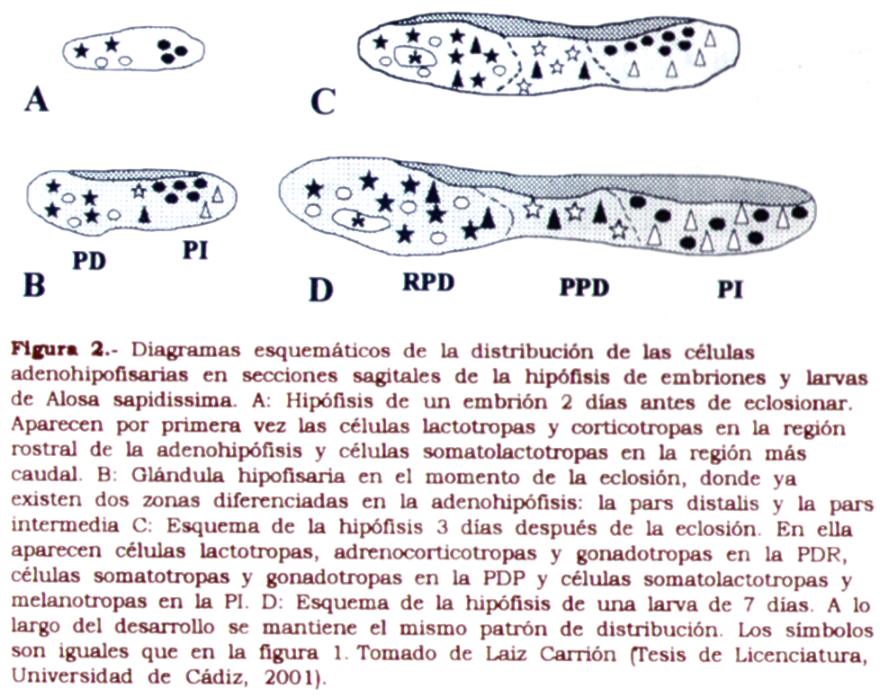
En peces teleósteos los procesos saltatorios existentes durante la ontogenia hacen necesario el estudio detallado del desarrollo hipofisario en cada especie. De este modo, el tiempo de aparición de las diferentes células adenohipofisarias varía con la especie estudiada [Balon, Am. Zool. 21: 573-596 (1981)]. Como regla general, las especies dulceacuícolas presentan un periodo de desarrollo embrionario mayor que las especies marinas. Durante el periodo embrionario los órganos endocrinos no son todavía funcionales, aunque se detectan diversas hormonas de origen materno. El papel fisiológico de las hormonas maternas no está claro, sin embargo su presencia en los huevos sugiere una importante función en el desarrollo y la supervivencia de los embriones [Tagawa e Hirano, J. Exp. Zool. 257: 360-366 (1991)]. La diferenciación del sistema endocrino generalmente ocurre antes de la absorción del saco vitelino. Este hecho sugiere que dicho sistema es importante para la transición de alimentación endógena a exógena. La diferenciación funcional de la glándula hipofisaria ocurre en un amplio rango de edades en las diferentes especies estudiadas, aunque siempre aparece asociada con procesos de desarrollo del ojo, justo antes del final de la absorción del saco vitelino [Tanaka et al., Aquaculture, 135: 111-126 (1995)]. En la figura 2 se muestra el momento de aparición de las distintas células adenohipofisarias en el clupeido Alosa sapidissima. Esta especie presenta un periodo embrionario de 6 días y es un buen modelo para el estudio del desarrollo adenohipofisario en teleósteos dulciacuícolas.
Implicaciones de las hormonas adeno-hipofisarias en el desarrollo
En teleósteos dulceacuícolas y eurihalinos la PRL es esencial para mantener el balance hidromineral en agua dulce. Sin embargo, en peces marinos la función de la PRL no está clara [Loretz y Bern, Neuroendocrinology, 35: 292-304 (1982)]. Así, la temprana aparición durante el desarrollo embrionario de inmunorreactividad para las células de PRL indican la existencia de células de PRL activas y sugieren una función de esta hormona en los procesos de desarrollo y crecimiento de embriones y larvas [Tanaka et al., Aquaculture. 135: 111-126 (1995)]. Además, la PRL ha sido relacionada con la respuesta al estrés y el momento de la eclosión supone un momento de estrés agudo para el animal. De este modo, la producción endógena de PRL antes de la eclosión podría servir para afrontar el estrés provocado por la misma y la posterior adaptación a un ambiente hipoosmótico.
La ACTH adenohipofisaria controla la síntesis del cortisol, que constituye el principal corticosteroide segregado por el tejido interrenal en peces. De este modo, deberíamos hablar de eje hipotálamo-hipofisario-interrenal con el cortisol como el producto final de secreción [Wendelaar Bonga, Physiol. Rev. 77: 591-625 (1997)]. El cortisol afecta a procesos de desarrollo en peces teleósteos, incluyendo la eclosión, el crecimiento y la metamorfosis [Yamano et al., J. Comp. Physiol. 161: 371-375 (1991)]. Así, es lógico que las células de ACTH sean unas de las primeras en presentar actividad en las etapas del desarrollo larvario, tanto en teleósteos marinos como dulceacuícolas.
La principal función de la GH es regular el crecimiento somático a través de la hipertrofia y/o la hiperplasia celular. Según la hipótesis dual, los efectos de GH se pueden ejercer por una acción directa de la hormona sobre un tejido o indirectamente mediante las somatomedinas o IGFs (Insulin-like growth factors) [Holly y Wass, J. Endocrinol. 122: 611-619 (1989)]. A nivel larvario, en teleósteos marinos no se ha encontrado inmunorreactividad en las células GH antes de la eclosión. El corto tiempo de desarrollo larvario que presentan los teleósteos marinos, así como el aporte materno de GH, podría explicar esta falta de actividad de las células productoras de GH. Sin embargo, los teleósteos dulciacuícolas, con un mayor periodo de desarrollo embrionario, sí presentan células GH activas antes de la eclosión [Tanaka et al., Aquaculture. 135: 111-126 (1995)]. Esto indica que la GH es ya importante en las primeras etapas del desarrollo ontogenético de peces teleósteos. Además, la producción local de IGFs en diferentes órganos del embrión es bastante importante, lo que puede favorecer los procesos de crecimiento sin necesidad de un gran aporte de GH.
Las GTHs son las hormonas que regulan la gametogénesis y la esteroidogénesis gonadal. En teleósteos se han descrito la existencia de dos GTHs: GTH I (GTH-vitelogénica) y GTH II (GTH II-maduracional) [Kah et al., Fish Physiol. Biochem. 11: 85-98 (1993)]. En diferentes especies de peces teleósteos no se ha detectado inmunorreactividad en las células secretoras de GTHs en estadios embrionarios. Durante el desarrollo larvario se ha puesto de manifiesto la expresión diferencial de ambas GTHs: las células conteniendo GTH I se detectan en las etapas larvarias coincidiendo con el momento en que tiene lugar la determinación morfológica gonadal, mientras que las células conteniendo GTH II no se detectan en ninguna etapa del desarrollo larvario [Saga et al., Gen. Comp. Endocrinol. 92: 233-241 (1993)]. Estos datos indican que la función de las GTHs no es necesaria hasta después de la eclosión.
El papel de la TSH es la estimulación de la glándula tiroides para la liberación de sus hormonas: tiroxina y triyodotiroxina. En peces teleósteos, estas hormonas están involucradas en procesos de metabolismo, osmorregulación y metamorfosis. Además, durante el desarrollo post-embrionario estimulan el crecimiento, actuando de un modo sinérgico con otras hormonas [Tagawa et al., Fish Physiol. Biochem. 8:515-520 (1990)]. En peces teleósteos se ha descrito inmunorreactividad en las células secretoras de TSH en los últimos estadios embrionarios o primeros estadios larvarios.
La
a-MSH presenta un papel en la adaptación al color de fondo mediante un control de la dispersión de los pigmentos en la piel. Sin embargo, debido a la relación estructural entre la á-MSH y la ACTH, también estimula la producción de cortisol y está involucrada en la respuesta al estrés [Lamers et al., J. Endocrinol. 135: 285-292 (1992)]. El papel de la á-MSH en el desarrollo larvario no está claro, aunque se relaciona con el proceso de estrés asociado a la eclosión. En la mayoría de las especies marinas las células melanotropas muestran inmunorreactividad después de la eclosión, mientras que en las especies dulceacuícolas se detecta dicha inmunorreactividad antes de la misma [Naito et al., Cell Tiss. Res. 272: 429-437 (1993); Saga et al., Anat. Embryol. 200: 469-475 (1999)].Las células secretoras de SL se activan por una extensa variedad de condiciones ambientales, aunque tanto en adultos como en larvas la función fisiológica de la SL es desconocida [Kaneko, Int. Rev. Cytol. 169: 1-24 (1996)]. Sin embargo, la pronta aparición de células sintetizadoras de SL en embriones sugiere un papel de esta hormona durante el desarrollo embrionario. De hecho, se ha propuesto un papel de la SL en la adaptación al estrés fisiológico asociado a la eclosión y al nuevo ambiente [Funkenstein et al., Fish Physiol. Biochem. 17: 247-252 (1997)].
Raúl Laiz Carrión es becario predoctoral en el Depto. de Biología, Facultad de Ciencias del Mar, Univ. de Cádiz. María del Pilar Martín del Río y Juan Miguel Mancera son Profesores Titulares en el Depto. de Biología, Facultad de Ciencias del Mar, Univ. de Cádiz.