Seres vivos: mucho más que un puñado de genes
Juan Carlos Aledo e Ignacio Fernández Molina
Vivimos un tiempo en el que los éxitos de la Biología Molecular son noticia. La oveja Dolly bien podría competir en popularidad con David Bisbal, y raro es el telediario en el que no se nos informa/desinforma sobre el hallazgo de un gen que nos permitirá curar no sé qué enfermedad. Los medios de comunicación han contribuido a difundir una falsa expectativa de las bondades de la ciencia moderna. Quizás por ello, no nos sorprende encontrar acalorados debates de cómo limitar en un futuro próximo los abusos de la biotecnología. ¿Sería o no ético manipular embriones para erradicar la violencia del comportamiento humano, o para crear individuos más inteligentes, etc.? Lo grave es que este injustificado optimismo haya calado en el seno académico. En un pasado curso de verano, un grupo de biólogos tuvimos oportunidad, como colofón al curso, de debatir sobre las limitaciones de la biotecnología. La opinión más extendida, y errónea a nuestro modesto entender, era que el fenotipo de un organismo se puede modificar a voluntad, sencillamente manipulando sus genes. Si se nos permite la ironía, diremos que posiblemente ello será así cuando los burros vuelen y las ranas críen pelos, ya sean estos burros y estas ranas transgénicos o no.
Si el objetivo de la biotecnología es modificar a voluntad el fenotipo de los organismos, tenemos que considerar que son muchos más, en número, los intentos fallidos que los que culminan con éxito. La capacidad de manipular genes es un hecho del presente, sin embargo, una cosa son los genes y otra bien distinta son los organismos. Esta distinción es fundamental y, sin embargo, no siempre se realiza. Aquí se plantea una paradoja: la biotecnología tiene como fin último modificar organismos y, sin embargo, muy frecuentemente se olvida de éstos como tales, centrando todo el interés en los genes de dichos organismos. Los genes, aún siendo importantes, no son más que meros instrumentos de los que se valen las leyes para actuar, y no debemos confundirlos con las propias leyes. En otros términos, la ecuación que define lo que es un organismo en un instante determinado, es una función multivariante dónde el genotipo no es la única, ni tan siquiera la principal, variable que determina el fenotipo.
Fenotipo = Genotipo + Ambiente + Historia + Azar
A nadie vamos a tener que convencer del papel que desempeñan el genotipo y el ambiente, y por tanto nos centraremos en algunos ejemplos que ilustran la importancia de los otros dos términos de esta ecuación: la historia y el azar.
Historia
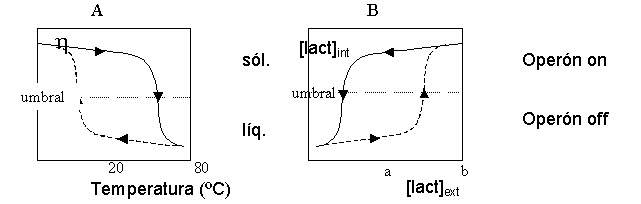
Aseguraba Borges que estamos hechos de tiempo. Podemos decir que el tiempo es un constituyente más de los sistemas biológicos, sobre todo cuando consideramos la vida como un proceso más que como una sustancia. Las leyes de la biología dependen de las leyes de la física y la química, pero también de una ingente cantidad de información sobre acontecimientos del pasado. Hay sistemas, tanto animados como inanimados, cuyo estado actual no está enteramente determinado por las condiciones actuales sino también por las pasadas. En otras palabras, la historia del sistema condiciona su presente y futuro. Imaginemos, por ejemplo, una suspensión de agar-agar. A 60ºC podemos encontrarlo líquido o sólido, dependiendo de su historia previa. Si el agar se calienta hasta los 100ºC y después se deja enfriar hasta los 60ºC, el estado de dicho agar será líquido. Si por el contrario, tomamos agar sólido a temperatura ambiente y vamos calentándolo hasta alcanzar los 60ºC, este agar permanecerá sólido (Ver figura 1A). Si el tiempo pasado es tan importante a la hora de determinar el estado de un sistema simple e inanimado como el agar, ¿qué influencia no tendrá en un ser vivo? Imaginemos ahora dos células de Escherichia coli genéticamente idénticas (igual genotipo), que se encuentran en el mismo medio (igual ambiente). Bien pudiera ser que ambas presenten, sin embargo, fenotipos muy distintos: una expresa los genes del operon lac mientras que la otra no; sencillamente porque la historia previa de ambas células es distinta. El operon lac contiene los genes que codifican para las proteínas implicadas en el metabolismo de la lactosa. Estos genes son expresados cuando una proteína reguladora que los reprime, deja de hacerlo al interaccionar con lactosa. Por tanto, la lactosa induce su propio metabolismo. Pero el operón posee una característica que enriquece el comportamiento de la bacteria frente a la lactosa, ya que uno de los genes del operón codifica para un transportador de lactosa.
Cuando hay lactosa en el medio, ésta entra y activa al operón, con lo que habrá más transportadores de lactosa y más lactosa intracelular con capacidad de activar el operón. Es un claro ejemplo de retroalimentación positiva. Así, aunque la concentración intracelular de lactosa necesaria para activar el operón es única, la concentración extracelular requerida para alcanzar dicha concentración intracelular es variable, dependiendo de la permeabilidad de la membrana (Fig. 1B). De esta forma, la permeabilidad de la membrana a la lactosa determina las concentraciones extracelulares umbrales necesarias para inducir o reprimir el operón, pudiendo ser muy diferentes según las circunstancias que hayan acompañado a la bacteria en el pasado (Fig. 1B). Así pues, en dos poblaciones genéticamente idénticas y en las mismas condiciones ambientales (igual concentración de lactosa en el medio), podemos encontrar células que están metabolizando lactosa y otras que no, debido a que las primeras estuvieron expuestas, en algún momento, a una concentración de lactosa en el medio suficientemente alta para inducir el operón, mientras que la otra no. Por este motivo, en esta población de bacterias la permeabilidad a la lactosa será alta, y la concentración de lactosa en el medio necesaria para mantener al operón inducido, será menor que para la otra población.
Azar
Incluso en una población de células clónicas (igual genotipo) que se encuentren en un mismo entorno (igual ambiente) y que aparentemente hayan tenido una misma historia, se pueden observar ligeras, y a veces no tan ligeras, variaciones morfológicas, distintas velocidades de desarrollo, y distintas concentraciones de las distintas biomoléculas presentes en cada célula (Proc. Natl. Acad. Sci. USA. 94: 814-819, 1997). Esta inevitable variabilidad fenotípica se fundamenta en la naturaleza estadística de las leyes fisicoquímicas (ver Encuentros en la Biología 57: 6-7). Esto es, cuando tenemos un sistema químico donde el número de moléculas reaccionantes es extremadamente bajo, como suele ser habitual en las células vivas, entonces las fluctuaciones aleatorias (ruido) son de magnitud considerable. De hecho, muchos circuitos genéticos emplean la redundancia, la retro-alimentación y otros artilugios para lograr una relación señal/ruido aceptable, cuando se requiere un funciona-miento determinista, es decir, no aleatorio (Trends Genet. 15: 65-59, 1999). No obstante, las células también han aprendido a explotar en su beneficio esta azarosa variabilidad. Son muchos y hetero-géneos los ejemplos conocidos que ilustran esta afirmación. El lector interesado puede dirigirse a la revisión (BioEssays 24: 512-518, 2002) donde se describen algunos de estos ejemplos. Aquí, valga un botón a modo de muestra. Aunque más que de botones vamos a hablar de pili. Las bacterias pueden expresar un número de apéndices externos llamados pili que facilitan la adhesión de la bacteria a determinadas superficies, como por ejemplo los órganos de los animales huésped. Por tanto, los pili son un determinante de virulencia de algunas bacterias patógenas. Un sistema bien estudiado es el que constituyen algunas cepas patógenas de E. coli. Que una de estas bacterias exhiba o no pili en su superficie depende del estado de activación de un operón llamado Pap (Pyelonephritis-associated pilus). Se ha comprobado que la activación/inactivación de este operón está estrictamente correlacionada con el estado de metilación de una región reguladora, que posee dos sitios de metilación, GATC1 y GATC2. La metilación de GATC2 (GmATC2) es necesaria para que se transcriba el operón (Fig. 2A). Hay una proteína reguladora, cuyo número por célula es bajo, que tiene una alta afinidad por los sitios GATC no metilados, de manera que se une a GATC1 o a GATC2 de forma aleatoria. Esta unión previene la metilación de dicho sitio. Si por azar la proteína reguladora se une a GATC2 protegiéndola de la metilación, se inhibe la transcripción del operón (Fig. 2B).
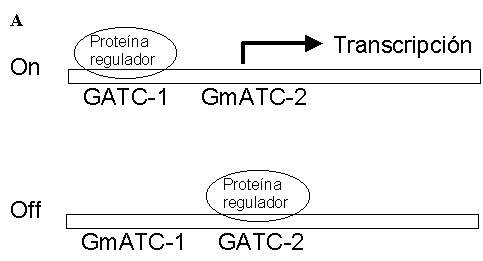
Figura 2. Regulación del operón Pap
En una población genéticamente homogénea y en condiciones ambientales idénticas, encontraremos bacterias que posean pili y otras que no, ya que cada individuo echa a cara o cruz la decisión de encender o apagar el operón responsable de la síntesis del pilus. No pretendemos ocultar que en ocasiones, la moneda utilizada puede estar trucada y que bajo determinadas condiciones ambientales las caras (digamos GmATC1) sean más frecuentes que las cruces (GmATC2); pero en cualquier caso, la expresión de pili es un proceso estocástico ya que aunque conozcamos el genotipo, el ambiente y la historia, nunca podremos precisar con grado de determinismo cuál será el fenotipo para un individuo dado. ¿Qué ventaja representa dejar al azar la expresión de pili? Para el individuo puede que ninguna, pero para la especie supone un enriquecimiento muy ventajoso. Los pili son estructuras altamente inmunogénicas, contar en todo momento con una subpoblación de bacterias que carezcan de estas estructuras, puede permitir evadir las defensas del huésped, para que más tarde, por azar, algunas de ellas vuelvan a desarrollar pili e infectar el tracto urinario.
Para finalizar nos gustaría hacer nuestras las palabras de Henrik Kacser, diciendo que cuando tratamos con seres vivos, el todo es mucho más que la suma de sus partes. Obstinarse en ignorarlo es condenarse al fracaso.
Juan Carlos Aledo es Profesor Titular de Bioquímica y Biología Molecular en la UMA.
Ignacio Fernández Molina es alumno de segundo ciclo en la UMA.